Genetyka
Genetyka
Genetyka to dziedzina biologii zajmująca się badaniem genów, zmienności genetycznej i dziedziczności w organizmach.
Chociaż dziedziczność obserwowano od tysiącleci, Gregor Mendel, naukowiec i zakonnik augustianów pracujący w XIX wieku, jako pierwszy nauczył się genetyki. Mendel badał „dziedziczenie cech”, wzorce w sposobie, w jaki cechy są przekazywane z rodziców na potomstwo. Zauważył, że organizmy (rośliny grochu) dziedziczą cechy poprzez odrębne „jednostki dziedziczenia”. Ten termin, wciąż używany do dziś, jest nieco niejednoznaczną definicją tego, co nazywa się genem.
Dziedziczenie cech i mechanizmy dziedziczenia molekularnego genów są nadal podstawowymi zasadami genetyki w XXI wieku, ale współczesna genetyka wykroczył poza dziedziczenie i zajął się badaniem funkcji i zachowania genów. Struktura i funkcja genów, zmienność i rozmieszczenie są badane w kontekście komórki, organizmu (np. Dominacja) oraz w kontekście populacji. Genetyka doprowadziła do powstania wielu dziedzin, w tym genetyki molekularnej, epigenetyki i genetyki populacyjnej. Organizmy badane w szerokim zakresie obejmują domeny życia (archeony, bakterie i eukarya).
Procesy genetyczne w połączeniu ze środowiskiem i doświadczeniami organizmu wpływają na rozwój i zachowanie, często określane jako natura kontra wychowanie. Wewnątrzkomórkowe lub zewnątrzkomórkowe środowisko żywej komórki lub organizmu może włączać lub wyłączać transkrypcję genów. Klasycznym przykładem są dwa nasiona identycznej genetycznie kukurydzy, jedno umieszczone w klimacie umiarkowanym, a drugie w klimacie suchym (bez wystarczającej ilości wodospadu lub deszczu). Podczas gdy średnią wysokość dwóch łodyg kukurydzy można określić genetycznie jako równą, ta w suchym klimacie rośnie tylko do połowy wysokości w klimacie umiarkowanym z powodu braku wody i składników odżywczych w jej środowisku.
Spis treści
Etymologia
Słowo genetyka wywodzi się ze starożytnej Grecji γενετικός genikos oznaczające „dopełniacz” / „generatywny”, które z kolei pochodzi od γένεσις geneza oznaczające „pochodzenie”.
Historia
Obserwacja, że żywe istoty dziedziczą cechy od swoich rodziców, była wykorzystywana od czasów prehistorycznych do ulepszania roślin uprawnych i zwierząt poprzez selektywną hodowlę. Współczesna genetyka, której celem jest zrozumienie tego procesu, rozpoczęła się w połowie XIX wieku wraz z pracą augustianina Gregora Mendla.
Przed Mendlem Imre Festetics, węgierski szlachcic, który mieszkał w Kőszeg przed Mendlem był pierwszym, który użył słowa „genetyka”. Opisał kilka zasad dziedziczenia genetycznego w swojej pracy Prawo genetyczne natury (Die genetische Gesätze der Natur, 1819). Jego drugie prawo jest takie samo, jak opublikowane przez Mendla. W swoim trzecim prawie rozwinął podstawowe zasady mutacji (można go uznać za prekursora Hugo de Vries).
Inne teorie dziedziczenia poprzedzały prace Mendla. Popularną teorią XIX wieku, zasugerowaną przez Charlesa Darwina w książce O pochodzeniu gatunków z 1859 r., Było mieszanie dziedziczenia: idea, że jednostki dziedziczą po rodzicach płynną mieszankę cech. Praca Mendla dostarczyła przykładów, w których cechy zdecydowanie nie były mieszane po hybrydyzacji, pokazując, że cechy są wytwarzane przez kombinacje różnych genów, a nie przez ciągłą mieszankę. Mieszanie cech u potomstwa jest teraz wyjaśnione działaniem wielu genów z efektami ilościowymi. Inną teorią, która miała wówczas pewne poparcie, było dziedziczenie cech nabytych: przekonanie, że jednostki dziedziczą cechy wzmocnione przez rodziców. Ta teoria (powszechnie kojarzona z Jean-Baptiste Lamarck) jest obecnie błędna - doświadczenia jednostek nie wpływają na geny, które przekazują swoim dzieciom, chociaż dowody z dziedziny epigenetyki ożywiły niektóre aspekty teorii Lamarcka. Inne teorie obejmowały pangenezę Karola Darwina (który miał zarówno nabyte, jak i odziedziczone aspekty) oraz przeformułowanie pangenezy przez Francisa Galtona jako cząstkowe i dziedziczne.
Genetyka mendlowska i klasyczna
Rozpoczęła się nowoczesna genetyka z badaniami Mendla nad naturą dziedziczenia roślin. W swoim artykule „ Versuche über Pflanzenhybriden ” („Eksperymenty nad hybrydyzacją roślin”), przedstawionym w 1865 r. W Naturforschender Verein (Society for Research in Nature) w Brünn, Mendel prześledził wzory dziedziczenia pewnych cech grochu i opisał je matematycznie. Chociaż ten wzorzec dziedziczenia można było zaobserwować tylko dla kilku cech, praca Mendla sugerowała, że dziedziczenie było cząstkowe, a nie nabyte, oraz że wzory dziedziczenia wielu cech można wyjaśnić za pomocą prostych reguł i stosunków.
Znaczenie pracy Mendla zyskało szerokie zrozumienie dopiero w 1900 roku, po jego śmierci, kiedy Hugo de Vries i inni naukowcy na nowo odkryli jego badania. William Bateson, zwolennik pracy Mendla, ukuł słowo genetyka w 1905 roku (przymiotnik genetyka , wywodzący się z greckiego słowa geneza —γένεσις, „pochodzenie” poprzedza rzeczownik i zostało po raz pierwszy użyte w sensie biologicznym w 1860 r.). Bateson zarówno działał jako mentor, jak i był w znacznym stopniu wspomagany przez pracę innych naukowców z Newnham College w Cambridge, w szczególności prace Becky Saunders, Nory Darwin Barlow i Muriel Wheldale Onslow. Bateson spopularyzował użycie słowa genetyka do opisania badań nad dziedziczeniem w swoim przemówieniu inauguracyjnym na Trzeciej Międzynarodowej Konferencji na temat Hybrydyzacji Roślin w Londynie w 1906 roku.
Po ponownym odkryciu metody Mendla prac naukowcy próbowali określić, które cząsteczki w komórce są odpowiedzialne za dziedziczenie. W 1900 roku Nettie Stevens zaczęła badać mącznika. W ciągu następnych 11 lat odkryła, że kobiety mają tylko chromosom X, a mężczyźni mają zarówno chromosomy X, jak i Y. Doszła do wniosku, że płeć jest czynnikiem chromosomalnym i jest determinowana przez mężczyznę. W 1911 roku Thomas Hunt Morgan argumentował, że geny znajdują się na chromosomach, na podstawie obserwacji mutacji białych oczu muszek owocówek związanych z płcią. W 1913 roku jego uczeń Alfred Sturtevant wykorzystał zjawisko powiązań genetycznych, aby pokazać, że geny są ułożone liniowo na chromosomie.
Genetyka molekularna
Chociaż geny były wiadomo, że istnieją na chromosomach, chromosomy składają się zarówno z białka, jak i DNA, a naukowcy nie wiedzieli, który z nich jest odpowiedzialny za dziedziczenie. W 1928 roku Frederick Griffith odkrył zjawisko transformacji (patrz eksperyment Griffitha): martwe bakterie mogą przenosić materiał genetyczny w celu „transformacji” innych wciąż żyjących bakterii. Szesnaście lat później, w 1944 roku, eksperyment Avery – MacLeod – McCarty zidentyfikował DNA jako cząsteczkę odpowiedzialną za transformację. Rola jądra jako repozytorium informacji genetycznej u eukariotów została ustalona przez Hämmerlinga w 1943 r. W jego pracy nad jednokomórkowym glonem Acetabularia . Eksperyment Hershey-Chase z 1952 roku potwierdził, że DNA (a nie białko) jest materiałem genetycznym wirusów infekujących bakterie, dostarczając dalszych dowodów na to, że DNA jest cząsteczką odpowiedzialną za dziedziczenie.
James Watson i Francis Crick określili strukturę DNA w 1953 r., korzystając z prac krystalografii rentgenowskiej Rosalind Franklin i Maurice'a Wilkinsa, które wskazały, że DNA ma strukturę helikalną (tj. ma kształt korkociągu). Ich model podwójnej helisy miał dwie nici DNA z nukleotydami skierowanymi do wewnątrz, z których każda pasowała do komplementarnego nukleotydu na drugiej nici, tworząc coś, co wygląda jak szczeble na skręconej drabinie. Ta struktura pokazała, że informacja genetyczna istnieje w sekwencji nukleotydów na każdej nici DNA. Struktura sugerowała również prostą metodę replikacji: jeśli nici są rozdzielone, można zrekonstruować nowe nici partnerskie dla każdej w oparciu o sekwencję starej nici. Ta właściwość nadaje DNA jego półkonserwatywny charakter, w którym jedna nić nowego DNA pochodzi z oryginalnej nici rodzicielskiej.
Chociaż struktura DNA pokazała, jak działa dziedziczenie, nadal nie było wiadomo, jak DNA wpływa na zachowanie komórek. W kolejnych latach naukowcy próbowali zrozumieć, w jaki sposób DNA kontroluje proces produkcji białek. Odkryto, że komórka wykorzystuje DNA jako szablon do tworzenia dopasowanego informacyjnego RNA, cząsteczek o nukleotydach bardzo podobnych do DNA. Sekwencja nukleotydowa informacyjnego RNA jest używana do tworzenia sekwencji aminokwasów w białku; ta translacja między sekwencjami nukleotydów a sekwencjami aminokwasów jest znana jako kod genetyczny.
Wraz z nowo odkrytym molekularnym zrozumieniem dziedziczenia nastąpiła eksplozja badań. Godna uwagi teoria powstała w 1973 roku od Tomoko Ohta wraz z jej poprawką do neutralnej teorii ewolucji molekularnej poprzez opublikowanie prawie neutralnej teorii ewolucji molekularnej. W tej teorii Ohta podkreślił znaczenie doboru naturalnego i środowiska dla tempa, w jakim zachodzi ewolucja genetyczna. Jednym z ważnych osiągnięć było sekwencjonowanie DNA końca łańcucha w 1977 roku przez Fredericka Sangera. Ta technologia umożliwia naukowcom odczyt sekwencji nukleotydów cząsteczki DNA. W 1983 roku Kary Banks Mullis opracował reakcję łańcuchową polimerazy, zapewniając szybki sposób izolowania i amplifikacji określonej części DNA z mieszaniny. Wysiłki Human Genome Project, Department of Energy, NIH i równoległe prywatne wysiłki Celera Genomics doprowadziły do sekwencjonowania ludzkiego genomu w 2003 roku.
Cechy dziedziczenia
Dyskretne dziedziczenie i prawa Mendla
Na najbardziej podstawowym poziomie dziedziczenie w organizmach zachodzi poprzez przekazywanie odrębnych dziedzicznych jednostek, zwanych genami, od rodziców do potomstwa. Właściwość tę po raz pierwszy zaobserwował Gregor Mendel, który badał segregację dziedzicznych cech grochu. W swoich eksperymentach badających cechy koloru kwiatów Mendel zauważył, że kwiaty każdej rośliny grochu były albo fioletowe, albo białe - ale nigdy nie były pośrednie między tymi dwoma kolorami. Te różne, odrębne wersje tego samego genu nazywane są allelami.
W przypadku grochu, który jest gatunkiem diploidalnym, każda roślina ma dwie kopie każdego genu, po jednej kopii odziedziczonej od każdego z rodziców. Wiele gatunków, w tym ludzie, ma ten wzór dziedziczenia. Organizmy diploidalne z dwiema kopiami tego samego allelu danego genu nazywane są homozygotami w tym locus genu, natomiast organizmy z dwoma różnymi allelami danego genu nazywane są heterozygotami.
Zbiór alleli dla danego organizmu nazywany jest jego genotypem, a obserwowalne cechy organizmu - jego fenotypem. Gdy organizmy są heterozygotyczne w genie, często jeden allel nazywany jest dominującym, ponieważ jego cechy dominują w fenotypie organizmu, podczas gdy inny allel nazywany jest recesywnym, ponieważ jego cechy zanikają i nie są obserwowane. Niektóre allele nie mają całkowitej dominacji i zamiast tego mają niepełną dominację poprzez wyrażanie fenotypu pośredniego lub kodominację poprzez wyrażanie obu alleli jednocześnie.
Kiedy para organizmów rozmnaża się płciowo, ich potomstwo losowo dziedziczy jeden z dwóch allele od każdego z rodziców. Te obserwacje dyskretnego dziedziczenia i segregacji alleli są zbiorczo znane jako pierwsze prawo Mendla lub prawo segregacji.
Notacja i diagramy
Genetycy używają diagramów i symboli do opisania dziedziczenia. Gen jest reprezentowany przez jedną lub kilka liter. Często symbol „+” jest używany do oznaczenia zwykłego, niezmutowanego allelu genu.
W eksperymentach z zapłodnieniem i hodowlą (a zwłaszcza przy omawianiu praw Mendla) rodzice są określani jako „P „pokolenie i potomstwo jako pokolenie„ F1 ”(pierwsze potomstwo). Kiedy potomstwo F1 kojarzy się ze sobą, potomstwo nazywane jest pokoleniem „F2” (drugie potomstwo). Jednym z typowych diagramów używanych do przewidywania wyniku krzyżowania jest kwadrat Punnetta.
Podczas badania chorób genetycznych u ludzi genetycy często używają wykresów rodowodowych, aby przedstawić dziedziczenie cech. Te wykresy odwzorowują dziedziczenie cechy w drzewie genealogicznym.
Wielokrotne interakcje genów
Organizmy mają tysiące genów, aw organizmach rozmnażających się płciowo geny te są generalnie sortowane niezależnie od siebie. Oznacza to, że dziedziczenie allelu dla koloru żółtego lub zielonego grochu nie jest związane z dziedziczeniem alleli dla białych lub fioletowych kwiatów. Zjawisko to, znane jako „drugie prawo Mendla” lub „prawo niezależnego asortymentu”, oznacza, że allele różnych genów są mieszane między rodzicami, tworząc potomstwo o wielu różnych kombinacjach. (Niektóre geny nie selekcjonują niezależnie, wykazując powiązania genetyczne, temat omówiony w dalszej części tego artykułu).
Często różne geny mogą oddziaływać w sposób wpływający na tę samą cechę. Na przykład u niebieskookiej Maryi ( Omphalodes verna ) istnieje gen z allelami określającymi kolor kwiatów: niebieski lub magenta. Jednak inny gen kontroluje, czy kwiaty mają w ogóle kolor, czy są białe. Kiedy roślina ma dwie kopie tego białego allelu, jej kwiaty są białe - niezależnie od tego, czy pierwszy gen ma allele niebieskie czy magenta. Ta interakcja między genami nazywa się epistazą, przy czym drugi gen jest epistatyczny do pierwszego.
Wiele cech nie jest cechami dyskretnymi (np. Fioletowe lub białe kwiaty), ale są cechami ciągłymi (np. Wzrost człowieka i kolor skóry) . Te złożone cechy są produktami wielu genów. Wpływ tych genów jest zależny w różnym stopniu od środowiska, którego doświadczył organizm. Stopień, w jakim geny organizmu wpływają na złożoną cechę, nazywany jest odziedziczalnością. Pomiar odziedziczalności cechy jest względny - w bardziej zmiennym środowisku środowisko ma większy wpływ na całkowitą zmienność cechy. Na przykład wzrost człowieka jest cechą o złożonych przyczynach. W Stanach Zjednoczonych odziedziczalność wynosi 89%. Jednak w Nigerii, gdzie ludzie mają bardziej zmienny dostęp do dobrego odżywiania i opieki zdrowotnej, wzrost jest dziedziczony tylko na 62%.
Molekularne podstawy dziedziczenia
DNA i chromosomy
Podstawą molekularną genów jest kwas dezoksyrybonukleinowy (DNA). DNA składa się z łańcucha nukleotydów, z których są cztery typy: adenina (A), cytozyna (C), guanina (G) i tymina (T). W sekwencji tych nukleotydów istnieje informacja genetyczna, a geny istnieją jako odcinki sekwencji wzdłuż łańcucha DNA. Wirusy są jedynym wyjątkiem od tej reguły - czasami wirusy używają bardzo podobnej cząsteczki RNA zamiast DNA jako materiału genetycznego. Wirusy nie mogą rozmnażać się bez żywiciela i nie mają na nie wpływu wiele procesów genetycznych, więc nie są uważane za żywe organizmy.
DNA normalnie istnieje jako dwuniciowa cząsteczka zwinięta w kształt podwójnej helisy. Każdy nukleotyd w DNA preferencyjnie łączy się w pary ze swoim nukleotydem partnerskim na przeciwległej nici: A tworzy pary z T, a C tworzy pary z G. Zatem w swojej dwuniciowej nić każda nić skutecznie zawiera wszystkie niezbędne informacje, zbędne z nicią partnerską. Ta struktura DNA jest fizyczną podstawą dziedziczenia: replikacja DNA powiela informację genetyczną poprzez dzielenie nici i używanie każdej nici jako szablonu do syntezy nowej nici partnerskiej.
Geny są ułożone liniowo wzdłuż długich łańcuchów sekwencji par zasad DNA. U bakterii każda komórka zwykle zawiera pojedynczy kolisty genofor, podczas gdy organizmy eukariotyczne (takie jak rośliny i zwierzęta) mają DNA ułożone w wiele liniowych chromosomów. Te nici DNA są często niezwykle długie; na przykład największy ludzki chromosom ma około 247 milionów par zasad. DNA chromosomu jest powiązane z białkami strukturalnymi, które organizują, zbierają i kontrolują dostęp do DNA, tworząc materiał zwany chromatyną; u eukariotów chromatyna składa się zwykle z nukleosomów, odcinków DNA owiniętych wokół rdzeni białek histonowych. Pełny zestaw dziedzicznego materiału w organizmie (zwykle połączone sekwencje DNA wszystkich chromosomów) nazywany jest genomem.
DNA najczęściej znajduje się w jądrze komórkowym, ale Ruth Sager pomogła w odkryciu geny nonchromosomalne znalezione poza jądrem. W roślinach często znajdują się one w chloroplastach i innych organizmach w mitochondriach. Te nonchromosomalne geny mogą być nadal przekazywane przez któregokolwiek z partnerów w rozmnażaniu płciowym i kontrolują różne cechy dziedziczne, które replikują się i pozostają aktywne przez pokolenia.
Chociaż organizmy haploidalne mają tylko jedną kopię każdego chromosomu, większość zwierząt a wiele roślin jest diploidalnych, zawierających po dwa z każdego chromosomu, a zatem po dwie kopie każdego genu. Dwa allele genu znajdują się na identycznych loci dwóch homologicznych chromosomów, każdy allel jest dziedziczony od innego rodzica.
Wiele gatunków ma tak zwane chromosomy płciowe, które określają płeć każdego organizmu. U ludzi i wielu innych zwierząt chromosom Y zawiera gen, który wyzwala rozwój specyficznie męskich cech. W ewolucji chromosom ten stracił większość swojej zawartości, a także większość genów, podczas gdy chromosom X jest podobny do innych chromosomów i zawiera wiele genów. To powiedziawszy, Mary Frances Lyon odkryła, że podczas reprodukcji dochodzi do inaktywacji chromosomu X, aby uniknąć przekazania dwukrotnie większej liczby genów potomstwu. Odkrycie Lyonu doprowadziło do odkrycia innych rzeczy, w tym chorób związanych z chromosomem X. Chromosomy X i Y tworzą silnie niejednorodną parę.
Reprodukcja
Kiedy komórki się dzielą, kopiowany jest ich pełny genom, a każda komórka potomna dziedziczy po jednej kopii. Proces ten, zwany mitozą, jest najprostszą formą rozmnażania i stanowi podstawę rozmnażania bezpłciowego. Rozmnażanie bezpłciowe może również wystąpić w organizmach wielokomórkowych, dając potomstwo, które dziedziczy genom od jednego rodzica. Potomstwo, które jest genetycznie identyczne z rodzicami, nazywane jest klonami.
Organizmy eukariotyczne często wykorzystują rozmnażanie płciowe do tworzenia potomstwa zawierającego mieszankę materiału genetycznego odziedziczonego po dwóch różnych rodzicach. Proces rozmnażania płciowego przebiega naprzemiennie między formami, które zawierają pojedyncze kopie genomu (haploid) i podwójne kopie (diploidalne). Komórki haploidalne łączą się i łączą materiał genetyczny, tworząc diploidalną komórkę ze sparowanymi chromosomami. Organizmy diploidalne tworzą haploidy, dzieląc się bez replikacji swojego DNA, tworząc komórki potomne, które losowo dziedziczą po jednym z każdej pary chromosomów. Większość zwierząt i wiele roślin jest diploidalnych przez większość swojego życia, a forma haploidalna jest zredukowana do jednokomórkowych gamet, takich jak plemniki lub jaja.
Chociaż bakterie nie używają haploidalnej / diploidalnej metody rozmnażania płciowego, bakterie mają wiele metod uzyskiwania nowych informacji genetycznych. Niektóre bakterie mogą ulec koniugacji, przenosząc mały okrągły fragment DNA do innej bakterii. Bakterie mogą również pobierać fragmenty surowego DNA znalezione w środowisku i integrować je ze swoim genomem - zjawisko to znane jest jako transformacja. Procesy te skutkują horyzontalnym transferem genów, przekazując fragmenty informacji genetycznej między organizmami, które w innym przypadku nie byłyby ze sobą powiązane. Naturalna transformacja bakteryjna zachodzi w wielu gatunkach bakterii i można ją traktować jako proces płciowy polegający na przenoszeniu DNA z jednej komórki do drugiej (zwykle tego samego gatunku). Transformacja wymaga działania wielu bakteryjnych produktów genowych, a jej podstawową funkcją adaptacyjną wydaje się naprawa uszkodzeń DNA w komórce biorcy.
Rekombinacja i powiązanie genetyczne
Diploidalna natura chromosomów umożliwia niezależne sortowanie genów na różnych chromosomach lub oddzielenie ich od pary homologicznej podczas rozmnażania płciowego, w którym powstają haploidalne gamety. W ten sposób u potomstwa pary godowej mogą pojawić się nowe kombinacje genów. Geny na tym samym chromosomie teoretycznie nigdy by się nie rekombinowały. Jednak robią to poprzez komórkowy proces krzyżowania chromosomów. Podczas krzyżowania chromosomy wymieniają odcinki DNA, skutecznie tasując allele genów między chromosomami. Ten proces krzyżowania się chromosomów zwykle występuje podczas mejozy, serii podziałów komórkowych, które tworzą komórki haploidalne. Wydaje się, że rekombinacja mejotyczna, szczególnie u drobnoustrojów eukariotycznych, pełni funkcję adaptacyjną naprawy uszkodzeń DNA.
Pierwszą cytologiczną demonstrację krzyżowania dokonały Harriet Creighton i Barbara McClintock w 1931 roku. Ich badania i eksperymenty kukurydza dostarczyła dowodów cytologicznych na teorię genetyczną, że połączone geny na sparowanych chromosomach w rzeczywistości wymieniają miejsca z jednego homologu na drugi.
Prawdopodobieństwo krzyżowania chromosomów między dwoma danymi punktami na chromosomie jest związane z odległość między punktami. W przypadku dowolnie długich dystansów prawdopodobieństwo krzyżowania się jest na tyle wysokie, że dziedziczenie genów jest skutecznie nieskorelowane. Jednak w przypadku genów, które są bliżej siebie, mniejsze prawdopodobieństwo krzyżowania oznacza, że geny wykazują powiązanie genetyczne; allele tych dwóch genów są zwykle dziedziczone razem. Wielkości powiązań między serią genów można łączyć, aby utworzyć liniową mapę powiązań, która z grubsza opisuje rozmieszczenie genów wzdłuż chromosomu.
Ekspresja genów
Kod genetyczny
Geny generalnie wyrażają swoje działanie funkcjonalne poprzez produkcję białek, które są złożonymi cząsteczkami odpowiedzialnymi za większość funkcji w komórce. Białka składają się z jednego lub więcej łańcuchów polipeptydowych, z których każdy składa się z sekwencji aminokwasów, a sekwencja DNA genu (poprzez pośrednik RNA) jest wykorzystywana do wytworzenia określonej sekwencji aminokwasowej. Proces ten rozpoczyna się od wytworzenia cząsteczki RNA z sekwencją pasującą do sekwencji DNA genu, w procesie zwanym transkrypcją.
Ta informacyjna cząsteczka RNA jest następnie wykorzystywana do wytworzenia odpowiedniej sekwencji aminokwasowej w procesie zwanym translacją . Każda grupa trzech nukleotydów w sekwencji, nazywana kodonem, odpowiada albo jednemu z dwudziestu możliwych aminokwasów w białku, albo instrukcji zakończenia sekwencji aminokwasowej; ta zgodność nazywana jest kodem genetycznym. Przepływ informacji jest jednokierunkowy: informacja jest przenoszona z sekwencji nukleotydów do sekwencji aminokwasowej białek, ale nigdy nie przenosi się z białka z powrotem do sekwencji DNA - zjawisko to Francis Crick nazwał centralnym dogmatem biologii molekularnej.
Specyficzna sekwencja aminokwasów skutkuje unikalną trójwymiarową strukturą tego białka, a trójwymiarowe struktury białek są powiązane z ich funkcjami. Niektóre są prostymi cząsteczkami strukturalnymi, jak włókna utworzone przez białko kolagenowe. Białka mogą wiązać się z innymi białkami i prostymi cząsteczkami, czasami działając jako enzymy, ułatwiając reakcje chemiczne w obrębie związanych cząsteczek (bez zmiany struktury samego białka). Struktura białka jest dynamiczna; białko hemoglobina wygina się w nieco inne formy, ponieważ ułatwia wychwytywanie, transport i uwalnianie cząsteczek tlenu we krwi ssaków.
Pojedyncza różnica nukleotydów w DNA może spowodować zmianę sekwencji aminokwasowej białka. Ponieważ struktury białek są wynikiem ich sekwencji aminokwasowych, niektóre zmiany mogą radykalnie zmienić właściwości białka poprzez destabilizację struktury lub zmianę powierzchni białka w sposób, który zmienia jego interakcję z innymi białkami i cząsteczkami. Na przykład anemia sierpowata jest ludzką chorobą genetyczną, która wynika z pojedynczej różnicy zasad w obrębie regionu kodującego część hemoglobiny β-globiny, powodującej zmianę pojedynczego aminokwasu, która zmienia właściwości fizyczne hemoglobiny. Sierpowate wersje hemoglobiny przyklejają się do siebie, tworząc włókna, które zniekształcają kształt czerwonych krwinek przenoszących białko. Te sierpowate komórki nie przepływają już płynnie przez naczynia krwionośne, mają tendencję do zatykania się lub degradacji, co powoduje problemy medyczne związane z tą chorobą.
Niektóre sekwencje DNA są transkrybowane na RNA, ale nie ulegają translacji na białko produkty - takie cząsteczki RNA nazywane są niekodującymi RNA. W niektórych przypadkach produkty te składają się w struktury, które są zaangażowane w krytyczne funkcje komórki (np. Rybosomalny RNA i transferowy RNA). RNA może również wywierać wpływ regulacyjny poprzez interakcje hybrydyzacyjne z innymi cząsteczkami RNA (np. MikroRNA).
Natura i wychowanie
Chociaż geny zawierają wszystkie informacje, których organizm używa do funkcjonowania, środowisko odgrywa ważną rolę w określaniu ostatecznych fenotypów, jakie wykazuje organizm. Wyrażenie „natura i wychowanie” odnosi się do tej uzupełniającej się relacji. Fenotyp organizmu zależy od interakcji genów i środowiska. Ciekawym przykładem jest umaszczenie kota syjamskiego. W tym przypadku temperatura ciała kota odgrywa rolę środowiska. Kocie geny kodują ciemną sierść, dlatego komórki produkujące sierść u kota wytwarzają białka komórkowe, co prowadzi do powstania ciemnych włosów. Ale te białka produkujące ciemne włosy są wrażliwe na temperaturę (tj. Mają mutację powodującą wrażliwość na temperaturę) i denaturują w środowiskach o wyższej temperaturze, nie wytwarzając pigmentu o ciemnej sierści w obszarach, w których kot ma wyższą temperaturę ciała. Jednak w środowisku o niskiej temperaturze struktura białka jest stabilna i normalnie wytwarza pigment o ciemnych włosach. Białko pozostaje funkcjonalne w obszarach skóry, które są zimniejsze - takich jak nogi, uszy, ogon i twarz - więc kot ma ciemne włosy na kończynach.
Środowisko odgrywa główną rolę w działaniu człowieka choroba genetyczna fenyloketonuria. Mutacja powodująca fenyloketonurię zaburza zdolność organizmu do rozkładania aminokwasu fenyloalaniny, powodując toksyczne nagromadzenie cząsteczki pośredniej, co z kolei powoduje poważne objawy postępującej niepełnosprawności intelektualnej i drgawek. Jeśli jednak ktoś z mutacją fenyloketonurii przestrzega ścisłej diety, w której unika się tego aminokwasu, pozostaje normalny i zdrowy.
Powszechna metoda określania, w jaki sposób geny i środowisko („natura i wychowanie”) przyczyniają się do fenotyp polega na badaniu bliźniąt jednojajowych i bliźniąt braterskich lub innego rodzeństwa urodzonego mnogiego. Identyczne rodzeństwo jest genetycznie takie samo, ponieważ pochodzi z tej samej zygoty. Tymczasem bliźnięta braterskie różnią się od siebie genetycznie tak samo, jak normalne rodzeństwo. Porównując, jak często określone zaburzenie występuje u pary bliźniąt jednojajowych z tym, jak często występuje u pary bliźniąt braterskich, naukowcy mogą określić, czy zaburzenie to jest spowodowane czynnikami środowiskowymi genetycznymi czy poporodowymi. Jeden ze słynnych przykładów obejmował badanie czworaczków genainowych, które były identycznymi czworaczkami, u których zdiagnozowano schizofrenię, jednak takie testy nie mogą oddzielić czynników genetycznych od czynników środowiskowych wpływających na rozwój płodu.
Regulacja genów
Genom danego organizmu zawiera tysiące genów, ale nie wszystkie te geny muszą być aktywne w danym momencie. Gen ulega ekspresji, gdy jest transkrybowany do mRNA i istnieje wiele komórkowych metod kontrolowania ekspresji genów, tak że białka są produkowane tylko wtedy, gdy są potrzebne komórce. Czynniki transkrypcyjne to białka regulatorowe, które wiążą się z DNA, promując lub hamując transkrypcję genu. Na przykład w genomie bakterii Escherichia coli istnieje szereg genów niezbędnych do syntezy aminokwasu tryptofanu. Jednak gdy tryptofan jest już dostępny dla komórki, te geny syntezy tryptofanu nie są już potrzebne. Obecność tryptofanu bezpośrednio wpływa na aktywność genów - cząsteczki tryptofanu wiążą się z represorem tryptofanu (czynnikiem transkrypcyjnym), zmieniając strukturę represora w taki sposób, że represor wiąże się z genami. Represor tryptofanu blokuje transkrypcję i ekspresję genów, tworząc w ten sposób regulację negatywnego sprzężenia zwrotnego w procesie syntezy tryptofanu.
Różnice w ekspresji genów są szczególnie wyraźne w organizmach wielokomórkowych, gdzie wszystkie komórki zawierają ten sam genom, ale mają bardzo różne struktury i zachowania ze względu na ekspresję różnych zestawów genów. Wszystkie komórki w organizmie wielokomórkowym wywodzą się z pojedynczej komórki, różnicując się na różne typy komórek w odpowiedzi na sygnały zewnętrzne i międzykomórkowe i stopniowo ustanawiając różne wzorce ekspresji genów, aby tworzyć różne zachowania. Ponieważ żaden pojedynczy gen nie jest odpowiedzialny za rozwój struktur w organizmach wielokomórkowych, wzorce te wynikają ze złożonych interakcji między wieloma komórkami.
W eukariotach istnieją strukturalne cechy chromatyny, które wpływają na transkrypcję genów, często w postaci modyfikacji DNA i chromatyny, które są stabilnie dziedziczone przez komórki potomne. Cechy te nazywane są „epigenetycznymi”, ponieważ istnieją „na wierzchu” sekwencji DNA i zachowują dziedziczenie z jednego pokolenia komórek do następnego. Ze względu na cechy epigenetyczne różne typy komórek hodowanych na tym samym podłożu mogą zachować bardzo różne właściwości. Chociaż cechy epigenetyczne są generalnie dynamiczne w trakcie rozwoju, niektóre, podobnie jak zjawisko paramutacji, mają wielopokoleniowe dziedziczenie i istnieją jako rzadkie wyjątki od ogólnej reguły DNA jako podstawy dziedziczenia.
Zmiana genetyczna
Mutacje
Podczas procesu replikacji DNA czasami zdarzają się błędy w polimeryzacji drugiej nici. Te błędy, zwane mutacjami, mogą wpływać na fenotyp organizmu, zwłaszcza jeśli występują w sekwencji kodującej białko genu. Wskaźniki błędów są zwykle bardzo niskie - 1 błąd na każde 10–100 milionów zasad - ze względu na zdolność polimerazy DNA do „korekty”. Procesy, które zwiększają tempo zmian w DNA, nazywane są mutagennymi: mutagenne substancje chemiczne sprzyjają błędom w replikacji DNA, często poprzez ingerowanie w strukturę parowania zasad, podczas gdy promieniowanie UV wywołuje mutacje, powodując uszkodzenie struktury DNA. Chemiczne uszkodzenie DNA również występuje naturalnie, a komórki wykorzystują mechanizmy naprawy DNA do naprawy niedopasowań i pęknięć. Naprawa nie zawsze jednak przywraca pierwotną kolejność. Wydaje się, że szczególnie ważnym źródłem uszkodzeń DNA są reaktywne formy tlenu wytwarzane przez komórkowe oddychanie tlenowe, które mogą prowadzić do mutacji.
W organizmach, które używają krzyżowania chromosomów do wymiany DNA i rekombinacji genów, błędy w wyrównaniu podczas mejoza może również powodować mutacje. Błędy w krzyżowaniu są szczególnie prawdopodobne, gdy podobne sekwencje powodują, że chromosomy partnerów przyjmują błędne dopasowanie; to sprawia, że niektóre regiony w genomach są bardziej podatne na mutacje w ten sposób. Błędy te powodują duże zmiany strukturalne w sekwencji DNA - duplikacje, inwersje, delecje całych regionów - lub przypadkową wymianę całych części sekwencji między różnymi chromosomami (translokacja chromosomów).
Naturalna selekcja i ewolucja
Mutacje zmieniają genotyp organizmu i czasami powodują pojawienie się różnych fenotypów. Większość mutacji ma niewielki wpływ na fenotyp organizmu, zdrowie lub sprawność reprodukcyjną. Mutacje, które mają wpływ, są zwykle szkodliwe, ale czasami niektóre mogą być korzystne. Badania na muchach Drosophila melanogaster sugerują, że jeśli mutacja zmienia białko wytwarzane przez gen, około 70 procent tych mutacji będzie szkodliwych, a pozostałe będą neutralne lub słabo korzystne.
Genetyka populacji bada rozmieszczenie różnic genetycznych w populacjach i jak te rozkłady zmieniają się w czasie. Na zmiany częstości występowania allelu w populacji wpływa głównie dobór naturalny, w którym dany allel zapewnia organizmowi selektywną lub reprodukcyjną przewagę, a także inne czynniki, takie jak mutacje, dryf genetyczny, autostop genetyczny, sztuczna selekcja i migracja .
Przez wiele pokoleń genomy organizmów mogą się znacznie zmieniać, co prowadzi do ewolucji. W procesie zwanym adaptacją, selekcja pod kątem korzystnych mutacji może spowodować ewolucję gatunku w formy lepiej zdolne do przetrwania w swoim środowisku. Nowe gatunki powstają w procesie specjacji, często powodowanym przez podziały geograficzne, które uniemożliwiają populacjom wymianę genów między sobą.
Porównując homologię między genomami różnych gatunków, można obliczyć odległość ewolucyjną między nimi oraz moment, w którym mogły się one rozdzielić. Porównania genetyczne są ogólnie uważane za dokładniejszą metodę charakteryzowania pokrewieństwa między gatunkami niż porównanie cech fenotypowych. Odległości ewolucyjne między gatunkami można wykorzystać do utworzenia drzew ewolucyjnych; drzewa te reprezentują wspólne pochodzenie i rozbieżność gatunków w czasie, chociaż nie wykazują transferu materiału genetycznego między niespokrewnionymi gatunkami (znanego jako poziomy transfer genów i najczęściej spotykany u bakterii).
Organizmy modelowe
Chociaż genetycy pierwotnie badali dziedziczenie w szerokim zakresie organizmów, naukowcy zaczęli specjalizować się w badaniu genetyki określonej podgrupy organizmów. Fakt, że dla danego organizmu istniały już znaczące badania, zachęciłby nowych badaczy do wybrania go do dalszych badań, a więc ostatecznie kilka organizmów modelowych stało się podstawą większości badań genetycznych. Typowe tematy badawcze w genetyce organizmów modelowych obejmują badanie regulacji genów i udziału genów w rozwoju i nowotworach.
Organizmy wybrano częściowo ze względu na wygodę - krótki czas pokolenia i łatwa manipulacja genetyczna sprawiły, że niektóre organizmy popularne narzędzia badawcze w dziedzinie genetyki. Powszechnie stosowane organizmy modelowe obejmują bakterię jelitową Escherichia coli , roślinę Arabidopsis thaliana , drożdże piekarskie ( Saccharomyces cerevisiae ), nicienie Caenorhabditis elegans , muszka owocówka ( Drosophila melanogaster ) i mysz domowa ( Mus musculus ).
Medycyna
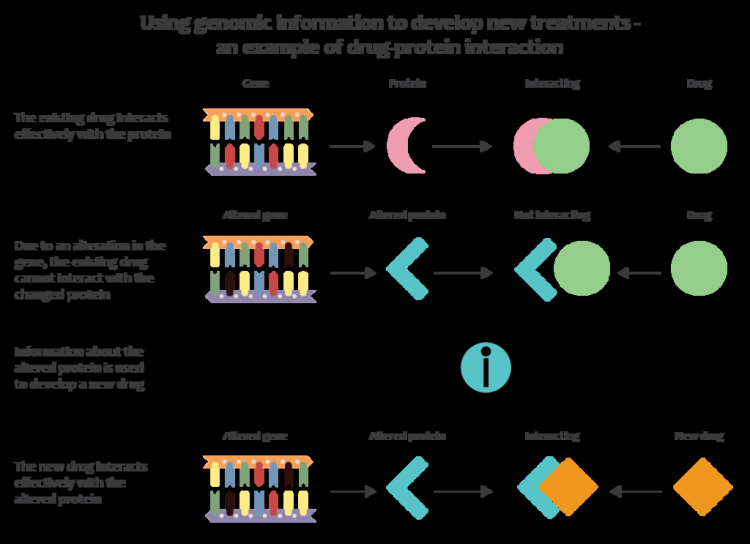
Genetyka medyczna stara się zrozumieć, w jaki sposób zmienność genetyczna wiąże się ze zdrowiem i chorobami człowieka. Szukając nieznanego genu, który może być zaangażowany w chorobę, naukowcy często używają powiązań genetycznych i wykresów rodowodu genetycznego, aby znaleźć lokalizację w genomie związanym z chorobą. Na poziomie populacji naukowcy wykorzystują randomizację mendlowską do poszukiwania lokalizacji w genomie, które są związane z chorobami, co jest metodą szczególnie przydatną w przypadku cech wielogenowych, które nie są jasno zdefiniowane przez pojedynczy gen. Po znalezieniu genu-kandydata często przeprowadza się dalsze badania nad odpowiadającymi mu (lub homologicznymi) genami organizmów modelowych. Oprócz badania chorób genetycznych, zwiększona dostępność metod genotypowania doprowadziła do powstania dziedziny farmakogenetyki: badania, w jaki sposób genotyp może wpływać na reakcje na leki.
Osoby różnią się dziedziczną skłonnością do zachorowania na raka i raka jest chorobą genetyczną. Proces rozwoju raka w organizmie to kombinacja zdarzeń. Mutacje czasami pojawiają się w komórkach ciała podczas ich podziału. Chociaż te mutacje nie zostaną odziedziczone przez żadne potomstwo, mogą wpływać na zachowanie komórek, czasami powodując ich wzrost i częstsze podziały. Istnieją mechanizmy biologiczne, które próbują zatrzymać ten proces; sygnały są wysyłane do nieprawidłowo dzielących się komórek, które powinny wywołać śmierć komórki, ale czasami pojawiają się dodatkowe mutacje, które powodują, że komórki ignorują te komunikaty. W organizmie zachodzi wewnętrzny proces naturalnej selekcji i ostatecznie mutacje gromadzą się w komórkach, aby promować ich własny wzrost, tworząc guz rakowy, który rośnie i atakuje różne tkanki ciała.
Zwykle komórka dzieli się tylko na odpowiedzi na sygnały zwane czynnikami wzrostu i przestaje rosnąć po kontakcie z otaczającymi komórkami oraz w odpowiedzi na sygnały hamujące wzrost. Zwykle dzieli się następnie określoną liczbę razy i umiera, pozostając w nabłonku, gdzie nie jest w stanie migrować do innych narządów. Aby stać się komórką rakową, komórka musi gromadzić mutacje w wielu genach (od trzech do siedmiu). Komórka rakowa może dzielić się bez czynnika wzrostu i ignoruje sygnały hamujące. Ponadto jest nieśmiertelny i może rosnąć w nieskończoność, nawet po kontakcie z sąsiednimi komórkami. Może uciec z nabłonka, a ostatecznie z guza pierwotnego. Następnie uciekająca komórka może przedostać się przez śródbłonek naczynia krwionośnego i zostać przetransportowana do krwiobiegu w celu skolonizowania nowego organu, tworząc śmiertelne przerzuty. Chociaż istnieją pewne predyspozycje genetyczne w niewielkiej części nowotworów, główna część jest spowodowana zestawem nowych mutacji genetycznych, które pierwotnie pojawiają się i gromadzą w jednej lub niewielkiej liczbie komórek, które dzielą się, tworząc guz i nie są potomstwo (mutacje somatyczne). Najczęstszymi mutacjami są utrata funkcji białka p53, supresora guza lub szlaku p53 i uzyskanie mutacji funkcji w białkach Ras lub innych onkogenach.
Metody badawcze
DNA można manipulować w laboratorium. Enzymy restrykcyjne są powszechnie stosowanymi enzymami, które tną DNA w określonych sekwencjach, tworząc przewidywalne fragmenty DNA. Fragmenty DNA można wizualizować za pomocą elektroforezy żelowej, która rozdziela fragmenty na podstawie ich długości.
Zastosowanie enzymów ligacyjnych umożliwia połączenie fragmentów DNA. Wiążąc („ligując”) fragmenty DNA z różnych źródeł, naukowcy mogą tworzyć rekombinowane DNA, DNA często kojarzone z organizmami zmodyfikowanymi genetycznie. Rekombinowane DNA jest powszechnie używane w kontekście plazmidów: krótkich kolistych cząsteczek DNA z kilkoma genami. W procesie znanym jako klonowanie molekularne, naukowcy mogą amplifikować fragmenty DNA poprzez wstawianie plazmidów do bakterii, a następnie hodowanie ich na płytkach agarowych (w celu wyizolowania klonów komórek bakterii - „klonowanie” może również odnosić się do różnych sposobów tworzenia klonowanych („ clonal ”).
DNA można również amplifikować stosując procedurę zwaną reakcją łańcuchową polimerazy (PCR). Używając określonych krótkich sekwencji DNA, PCR może izolować i wykładniczo amplifikować docelowy region DNA. Ponieważ może amplifikować z bardzo małych ilości DNA, PCR jest również często używany do wykrywania obecności określonych sekwencji DNA.
Sekwencjonowanie DNA i genomika
Sekwencjonowanie DNA, jedno z najbardziej podstawowe technologie opracowane w celu badania genetyki, pozwalają badaczom określić sekwencję nukleotydów we fragmentach DNA. Technika sekwencjonowania terminacji łańcucha, opracowana w 1977 roku przez zespół kierowany przez Fredericka Sangera, jest nadal rutynowo stosowana do sekwencjonowania fragmentów DNA. Korzystając z tej technologii, naukowcy byli w stanie zbadać sekwencje molekularne związane z wieloma chorobami człowieka.
Ponieważ sekwencjonowanie stało się tańsze, naukowcy zsekwencjonowali genomy wielu organizmów za pomocą procesu zwanego składaniem genomu, który wykorzystuje narzędzia obliczeniowe do łączenia sekwencji z wielu różnych fragmentów. Technologie te zostały wykorzystane do sekwencjonowania ludzkiego genomu w projekcie Human Genome Project zakończonym w 2003 r. Nowe technologie sekwencjonowania o wysokiej przepustowości radykalnie obniżają koszt sekwencjonowania DNA, a wielu badaczy ma nadzieję, że koszt ponownego sekwencjonowania ludzkiego genomu spadnie do tysiąca dolarów.
Sekwencjonowanie nowej generacji (lub sekwencjonowanie o wysokiej przepustowości) powstało z powodu stale rosnącego zapotrzebowania na sekwencjonowanie o niskich kosztach. Te technologie sekwencjonowania umożliwiają jednoczesne wytwarzanie potencjalnie milionów sekwencji. Duża ilość dostępnych danych sekwencyjnych stworzyła dziedzinę genomiki, badań wykorzystujących narzędzia obliczeniowe do wyszukiwania i analizowania wzorców w pełnych genomach organizmów. Genomikę można również uznać za poddziedzinę bioinformatyki, która wykorzystuje podejścia obliczeniowe do analizowania dużych zbiorów danych biologicznych. Częstym problemem w tych dziedzinach badań jest zarządzanie i udostępnianie danych dotyczących ludzi i informacji umożliwiających identyfikację osoby.
Społeczeństwo i kultura
19 marca 2015 r. Grupa czołowi biolodzy zaapelowali o ogólnoświatowy zakaz klinicznego stosowania metod, w szczególności CRISPR i palca cynkowego, w celu edycji ludzkiego genomu w sposób, który może być dziedziczony. W kwietniu 2015 r. Chińscy naukowcy opublikowali wyniki podstawowych badań dotyczących edycji DNA nieżywotnych embrionów ludzkich przy użyciu CRISPR.
Gugi Health: Improve your health, one day at a time!

Gen pamięci może napędzać PTSD
Żywe wspomnienia mogą być atutem, jeśli uczysz się do egzaminu lub próbujesz …

Genetyka może nie mieć wiele wspólnego z przewidywaną długością życia, ale tak właśnie jest
Według nowych badań to, jak długo żyjemy, wydaje się mieć mniej wspólnego z …